
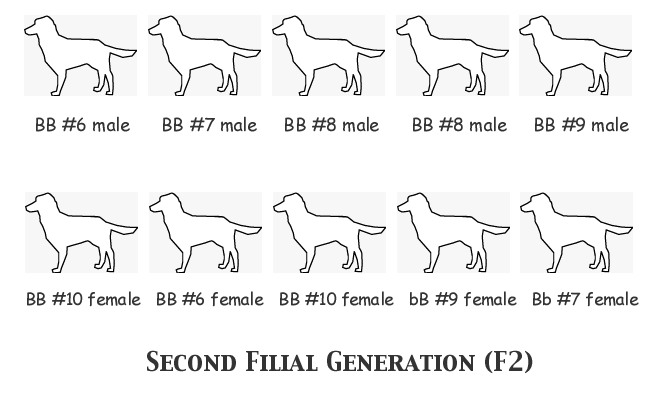
This time mother nature threw the breeder a curve, as she experienced that not-uncommon and sometimes hoped-for phenomenon -- an all-male litter -- in the #8 mating. Since div> her plan has been to select two pups from each litter to carry on the breeding programme, she had to select two males from Litter #8 and compensate by taking two females from Litter #10. As luck would have it, she has unknowingly selected five BB genotype males! The lone liver-nosed female was not selected for breeding; she had a lot of competition in her litter and it may be that the breeder just liked one of the black-nosed heterozygote sisters better, we'll never know.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
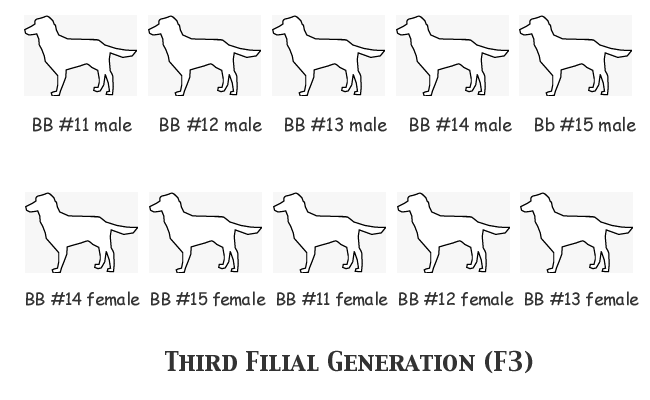
On we go to the third filial generation of this experimental breeding colony. This time we see five nice litters of six puppies each. Three of those litters could not produce anything but the BB genotype, since both parents were BB. The other two matings were both homozygote BB males to heterozygote bitches. Of those two, one conformed perfectly to theoretical expectations, while the other (#14) produced only two heterozygote puppies instead of the theoretical three. There is nothing in any way remarkable in any of these litters; yet the gene frequency has shifted again, such that this time the allele frequencies are FB = 0.9167, Fb = 0.0833. There are now so few carriers of the recessive b gene in this population -- five pups out of thirty -- that chance must now play an increasingly large part in what happens next.
From three of the five litters the breeder will inevitably pick BB males and females. In litter #14 there are equal numbers of BB and Bb males and the odds are even that she'll pick a homozygote; that is what happens in the event. In Litter #15 it's a similar situation, and this time by chance she picks a heterozygote male! (If by chance she had chosen another homozygote, the recessive b gene would have been lost in the next generation.) Litter #14 has only BB females, so that is what is chosen. In Litter #15 it's down to pick with one female of each genotype; as it happens, the BB girl gets chosen. The allele frequencies of the breeding population turn out to be FB = 0.95 and Fb = 0.05 (if the other female had been chosen the ratio would have been 90/10). It should be obvious that by this point there is really only one way the situation can ultimately turn out, but let's go ahead and tabulate the litters of the fourth filial generation.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Out of thirty puppies in this generation, only two are heterozygotes. As sheer blind luck would have it, the recessive allele will not disappear with this generation, because it is certain that one of the two heterozygote females from Litter #20 must be chosen; yet the situation will not improve in the next generation, unless by chance there are both male and female heterozygotes in the litter bred from that female, and unless one of each is chosen to represent that litter. The odds are against that happening. So chances are the next generation will either repeat the situation shown in this F4 generation, or else the recessive will finally be lost.
I hope that the reader has paid some attention to the distribution of males and females in these twenty litters. Of a total of 118 puppies whelped, half were males and half were females, as one would expect in any large group of progeny. Obviously the theoretical expectation is 50/50 -- yet, as any experienced breeder well knows, the sex ratio in any particular litter can be anything at all. Indeed, this variability of the sex ratio in each litter, seen against the background of an iron-clad 50/50 expectation, is the strongest possible demonstration of the effect of sampling error in binomial sampling with replacement, when only small samples are taken. If so much variability can show up in individual litters with respect to the trait of male or female sex, then obviously we can expect errors just as great in other genetic traits, just as much in those we never notice as in those we track closely.
I have played quite fairly with the reader in this example, yet I have effectively demonstrated how, within a small breeding population such as might be maintained by almost any breeder, a recessive allele with an initial incidence of thirty percent can be lost from the population within five or six generations -- solely through the operation of genetic drift amplified by normal selection practices. It is important to realise that the recessive was actively selected against only once, if then -- when the breeder failed to choose the liver-nosed female in Litter #9; and in fact that pup may have been passed over for reasons that had nothing to do with the colour of her nose. What is significant is that in each generation ten animals were selected out of a population of twenty-five to thirty-two. There is nothing unusual about this level of selection; it would be normal in most small or medium-sized kennels. Yet all it takes is sampling error plus this level of selection -- not even selection directed at the B-locus trait, just undifferentiated selection -- to push the original 30% recessive into oblivion.
Obviously the example I have just set forth is not the only possible course that such a situation can take; things can conceivably go the other way. The fact that the gene eliminated was a recessive one has little significance unless we are talking about traits that are obvious to gross examination. It IS more difficult to lose a dominant gene that can readily be seen, simply because the whole process takes place in plain view, because every individual with a single copy of a dominant colour gene (for example) shows his possession of that gene in his phenotype. But if we are talking about traits that cannot be seen by the eye, then minority dominant genes disappear just as readily as recessives. I believe that this is what has actually happened in many purebred registered dog breeds: dominant genes have disappeared from small breeding populations through drift plus selection, selection that was directed towards recessive traits of appearance found desirable in the show ring, leaving those breeds with homozygous recessive genotypes for many of the "genetic defects" that show up with such alarming persistence in today's purebred dog populations.
To finish reading "Random Drift -- The Breeder's Hidden Enemy" click here.